




Molecular details of GPCRs, their G protein coupling and arrestin interactions are providing unprecedented insight into signalling cascades and are often achieved through judicious antibody stabilization or protein engineering1. Meanwhile, recognition of the importance of lipids in mediating GPCR signalling interactions has been derived from the use of lipid nanodiscs and peptidiscs to recreate membrane environments5,6,7. Capturing signalling of unmodified GPCRs, in native membrane environments, has so far eluded biophysical measurement. Here, selecting the best-characterized GPCR rhodopsin, the dim light receptor of the mammalian visual system8, we formed lipid vesicles from disc membranes of dark-adapted bovine retinal rod outer segments (ROSs) and ejected this GPCR directly from fragments of its native membrane environment into a mass spectrometer (Fig. 1a–c, Supplementary Video 1). We monitored signalling by operating our mass spectrometer in a dark room and controlling light exposure to prompt photoisomerization of 11-cis-retinylidene (cis-retinal rho) to form activated all-trans-retinylidene (rho*). The Schiff base of rho* was then hydrolysed to the apo-protein opsin and all-trans-retinal9,10. During photoactivation, rho* interacted with GDP-bound transducin (heterotrimeric G protein (Gt)), leading to GDP–GTP exchange and subsequent dissociation and formation of Gαt•GTP, which then interacted with phosphodiesterase 6 (PDE6), displacing the PDE6 γ-subunit, releasing inhibition of catalysis and resulting in rapid hydrolysis of cytoplasmic cGMP11. Here we show that all components of this signalling pathway (protein, downstream effectors, cofactors and lipids) can be ejected simultaneously and their response to photon activation in the native membrane captured by native mass spectrometry directly and in real time (Fig. 1d).

a, Following absorption of a photon of light hv (1), 11-cis-retinal of rho isomerizes to the all-trans isomer (2). The activated states of rho (rho*) engage with transducin (Gt), consisting of Gαt•GDPβγt, and exchange GDP for GTP. Gt dissociates to form Gαt•GTP and Gβγt; loss of retinal from rho* leads to the formation of opsin (3). α-Subunits of Gt interact with the γ-subunits in the PDE6 enzyme, with γ-subunits undergoing a conformational change, relieving inhibition and thereby activating PDE6 to cause hydrolysis of cGMP (4). Depletion of cGMP then closes the ion channel and the ‘dark current’ is terminated (5). The resulting change in the membrane potential produces the sensation of light. b, Disc membranes of rod cells are homogenized to form a heterogeneous distribution of vesicles that are introduced directly into the mass spectrometer (MS). The spectrum shown was recorded using the parameters stated above, which led to the dissociation of lipids from proteins. c, An LED light source is configured to apply timed light intervals before the electrospray ionization of the vesicles into an Orbitrap mass spectrometer under the conditions noted. HCD, higher-energy collisional dissociation. d, Following the addition of a soluble fraction containing PDE6 and Gt, all proteins along the signalling pathway were detected. The proteins were ejected intact as rho/opsin (red), trimeric Gt•GDP (cyan) and tetrameric PDE6•cGMP (pink). The mass spectrum shown from m/z 4,000 to m/z 8,000 represents the raw data. The experiment was repeated three times.
To establish mass spectrometry and light conditions that enable monitoring of rhodopsin signalling, and before instituting our approach in native membranes, we used membrane solubilization and detergents to extract rhodopsin from dark-adapted ROS membrane, as previously described for analysis via mass spectrometry12,13,14. Following extraction into lauryl maltose neopentyl glycol (LMNG), we identified illumination conditions (LED-emitting cold white light; Extended Data Fig. 1) and mass spectrometry conditions to release rho from membrane lipids and detergents and to monitor rho* signalling through the mass change associated with conversion of rho to opsin (Δ mass = 266 Da). Before illumination, 73 ± 1% rho (11-cis-retinal or trans-retinal) and 27 ± 1% opsin were present. Recording of spectra in real time was synchronized with illumination; equal population of both species was achieved after 3 min (Fig. 2a), reaching a plateau with 27 ± 1% rho remaining at 20 min in this detergent micelle preparation (Fig. 2c). We next established mass spectrometry conditions that enabled ejection of rho and opsin (rho/opsin) directly from disc membrane fragments while activated with light. To do this, we adapted and applied a sonicated lipid vesicle approach15,16 to purified ROS disc membranes, forming vesicles for direct introduction into the mass spectrometry. Under these conditions, monomeric rho was ejected. No dimeric population was observed, attributed to the relatively small dimer interface observed via cryo-electron microscopy in nanodiscs17 that dissociates under these mass spectrometry conditions. Comparing the post-translational modification status of dark-adapted and light-exposed rho/opsin extracted in detergent or ejected from membranes, we found that glycosylation, palmitoylation and cysteinylation status are largely indistinguishable (Extended Data Fig. 2a–d, Extended Data Table 1). An increase in phosphorylation of opsin was observed 20 min after signalling was initiated, both in detergent-extracted and membrane-ejected rho/opsin, consistent with progress towards termination of this signalling pathway through eventual interaction with arrestin18.
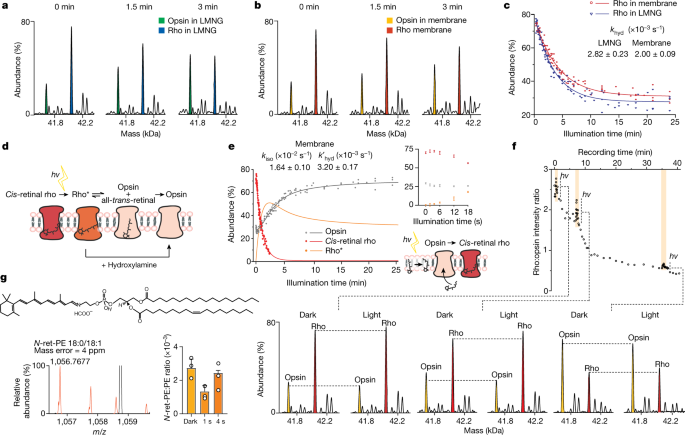
a, b, Changes in the population of rho and opsin in LMNG (blue and green, respectively; a) and in native membranes (red and yellow, respectively; b). Individual spectra are shown as zero-charge plots with illumination times stated. c, Plot of the relative abundance of rho during illumination, in either LMNG detergent micelles (blue) or native membranes (red) monitored as a change in mass as a function of time. Rate constants for hydrolysis are shown for the reaction in detergent micelles or in membranes (Extended Data Fig. 3). d, Schematic of the dark-adapted state undergoing light-activated conversion of cis-retinal to all-trans-retinal, hydrolysis of the Schiff base and dissociation from rho to form opsin. e, Monitoring the decay of cis-retinal rho in membranes pretreated with hydroxylamine (red), the generation of opsin in native membranes (grey) and the change of rho* calculated over the illumination period (orange). The inset, which is an expansion of the illumination period from 0–18 s, shows a less than 18-s increase in rho abundance at the expense of opsins. The schematic depicts a possible regeneration mechanism through isomerization of trans-retinal. f, Monitoring the conversion of rho to opsin and the regenerative reaction of opsin to rho following pulses of light at 0.51 min, 7 min and 35 min for time periods of 3 s, 12 s and 32 s, respectively. Zero-charge mass spectra are shown at time points during this conversion reaction, before illumination (dark) and at the peak of the three light pulses. g, Spectrum (orange) of N-ret-PE (18:0/18:1 representative structure shown) extracted from native membranes; the bar graph shows changes in the equilibrium between conjugated and free PE in response to illumination time. Data are presented as mean ± s.d. (n = 3).
Monitoring rho/opsin populations during continuous light exposure in membrane vesicle fragments, we found that after 3 min rho remained predominant over opsin (Fig. 2b). The reaction in membranes proceeds until a plateau is reached with approximately 32% rho remaining after 20 min (compared with approximately 27% in detergent micelles after the same time period (Fig. 2c)). We next calculated and compared the rates of chromophore hydrolysis (khyd; at pH 7.0 and 28 °C) in LMNG micelles and from ROS membrane vesicles according to a kinetic model (Fig. 2c, Extended Data Fig. 3, eq. 7). As the hydrolysis of all-trans-retinal from rho* is a slower step than photoisomerization of 11-cis-retinal to trans-retinal in rho, our results allow us to conclude that hydrolysis of all-trans-retinylidene from rho* is slower in vesicles than in detergent micelles. Considering the isomerization reaction, loss of all-trans-retinal from the binding pocket of rho* is monitored through a change in mass following hydrolysis of the Schiff base. The extent of cis-retinal or trans-retinal bound to opsin before hydrolysis, however, is not apparent via mass alone. To delineate these isomeric populations, we adopted a well-established protocol in which ROS membrane preparations were pre-incubated in the dark with 5 mM hydroxylamine (Fig. 2d). Rho in its ground state is insensitive to hydroxylamine but, upon photoisomerization of cis-retinal to all-trans-retinal, hydroxylamine is able to attack and cleave the Schiff base in the photoactivated rhodopsin to form all-trans-retinyl oxime19. As expected, rho* in membranes decayed significantly faster in the presence of hydroxylamine (Extended Data Fig. 2e). This hydroxylamine experiment, together with the rho/opsin photoconversion of untreated membranes mentioned above (khyd), enabled us to subtract the two decay curves and consequently obtain the rates of retinal isomerization (kiso) and hydrolysis without isomerization (k′hyd) in native membranes (Fig. 2e, Extended Data Fig. 3).
Careful examination of the data reveals an initial phase in which opsin regenerates to rho (less than 18 s) in competition with the overall rho/opsin conversion. The increase in cis-retinal rho at 2.7 ± 1 s is coincident with the lowest level of opsin at 3.6 ± 1 s (Fig. 2e, inset). This initial reaction, in the absence of hydroxylamine, must therefore arise primarily from cis-retinal binding to opsin. To determine whether this observation was a feature of the membrane, we applied short-term illumination on disc membrane preparation and to rho purified in LMNG detergent micelles. Upon each transient illumination of rho from ROS, a rapid rise of rho was observed together with the general trend of rho transforming to opsin (Fig. 2f). Some evidence for regeneration of rho in micelles was observed, although to a much lower extent (Extended Data Fig. 4). The ability to regenerate is a known property of the visual system, which requires the conversion of all-trans-retinal to 11-cis-retinal and has been shown to be carried out by enzyme pathways in neighbouring cells10. Our proteomics experiments (Supplementary Table 1) were unable to detect key enzymes from the regeneration cycle, such as retinal GPCR (RGR) or the retinoid isomerase RPE65 (ref. 20), prompting us to consider the possibility that this rapid regeneration (less than 18 s) may be a feature of the membrane itself.
Previous reports have implicated an N-all-trans-retinyl-PE conjugate that can undergo photoisomerization to form cis-retinals (primarily 11-cis-retinal (85–86%) with small contributions from 9-cis-retinal (12–13%) and 13-cis-retinal (1–2%))21. Although our mass spectrometry approach cannot readily distinguish these structural isomers, our lipidomics data reveal the presence of high concentrations of polyunsaturated fatty acids including PE (Extended Data Fig. 5a–c). To explore the formation of retinal conjugated to PE, we extracted lipids into isopropanol directly from dark-exposed and controlled-light-exposed ROS disc membranes. We selected three representative lipids (PE 18:0/18:1, 18:0/20:4 and 18:0/22:5) with minimal overlap in mass spectra, and searched for their respective retinyl conjugation. We found direct evidence for the formation of N-retinyl-PE (N-ret-PE) in all cases and monitored its change as a function of exposure to light. After 1 s of illumination, a significant decrease in conjugation was observed (more than 2.5-fold for PE 18:0/18:1), consistent with light-stimulated retinal dissociation from PE (Fig. 2g, Extended Data Fig. 5d–f). Peaks assigned to N-ret-PE increased after 4 s of light exposure to similar levels as the dark-adapted membrane, suggesting further conjugation events. Significantly reduced rho regeneration was observed for rho purified in micelles, where a lack of the conjugation system for retinal and PE would be anticipated (Extended Data Fig. 4). Moreover, regeneration of rho might be expected to be limited by the availability of cis-retinal. To investigate this possibility, we incubated intact ROS disc membranes with a tenfold excess of all-trans-retinal. Following illumination, regeneration of rho from 79% to 91% was observed after 3.8 min (Extended Data Fig. 6). Together, these results imply that conversion of opsin to rho in membranes can be supplemented by the addition and photoisomerization of all-trans-retinal and that conjugation of PE provides a source of cis-retinal for regeneration in membranes, a mechanism that has been suggested for sustained vision in daylight21.
Turning our attention to the next step in the signalling cascade, rho* signals through Gt, releasing GDP and dissociating to form Gαt•GTP for interaction with PDE6. Before illumination, GDP-bound heterotrimeric Gt•GDP and cGMP-bound heterotetrameric PDE6 (PDE6•cGMP) were observed as the predominant species (Fig. 1d). To monitor real-time signalling through transducin, excess soluble fraction containing Gt and PDE6 was added to our membrane preparation (see Methods). We monitored the decrease in the ratio of Gt•GDP:Gt as a function of time, initially for periods of up to 1 min (Fig. 3c), but found little change beyond the first 15 s of light stimulation. The spectra of Gt are complicated by the presence of additional proteoforms (α2 from cone Gt (Extended Data Fig. 7a–c)). Control experiments in the absence of light or ROS membranes (Extended Data Fig. 7d–f) confirm that light activation of rho in the membrane is required to decrease the population of Gt•GDP during signalling (Fig. 3a).
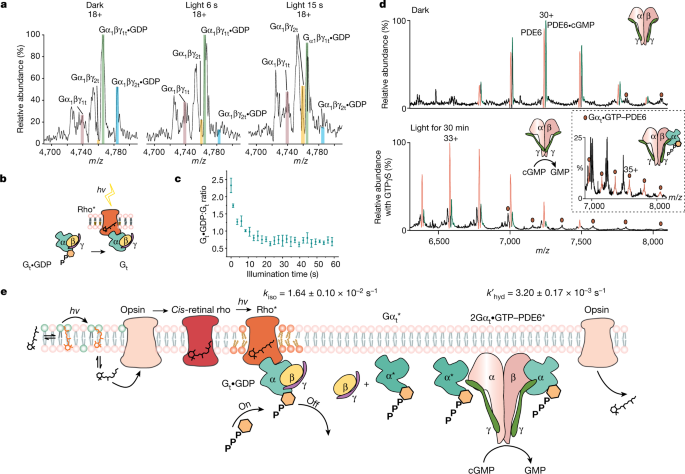
a, Native mass spectra of Gt•GDP with no additional GTP added containing two α-isoforms (α1 and α2) and two γ-isoforms (γ1 and γ2) are catalysed to apo-Gt during a 15-s time course of illumination. b, Reaction scheme: rho* catalysed the nucleotide exchange of Gt. c, The ratio of ground-state Gt•GDP to intermediate apo-Gt decreases rapidly during the first 15 s after illumination. Data are presented as mean ± s.e. (n = 3). d, Native mass spectra of PDE6 ejected from ROS disc membrane vesicles under dark conditions reveal that cGMP binds to intact tetrameric PDE6 with an approximately 1:1 ratio (top). A low population of PDE6–Gαt•GTP is also observed (red circles). After exposure to light in the presence of ROS disc membranes, the population of PDE6 bound to cGMP is reduced considerably, consistent with the release of GMP following its hydrolysis via activated PDE6 with addition of a molar equivalent of GTPγS to Gt (bottom). e, Schematic shows the rho* signalling cascade that involves the conjugation of all-trans-retinal with PE and its light activation to form cis-retinal that is able to interact with opsin. Light-activated conversion of rho to an intermediate state takes place with rho* capable of interacting with Gt. Changes in the lipid bilayer are depicted as unsaturated lipids that are recruited during rho* signalling (lipids, orange). The Gαt•GTP subunit produced following hydrolysis of Gαt•GDP interacts with PDE6, relieving its inhibition via the γ-subunit and effecting the hydrolysis of cGMP, which is then released from PDE6. Gαt•GDP is formed for the regeneration of Gt•GDP. All experiments shown in this figure were repeated at least three times. Data are presented as mean ± s.e. (n = 3).
To explore the relationship between Gt-dependent rho* signalling and PDE6•cGMP hydrolysis, we investigated three different GTP conditions: (1) with endogenous levels of GTP in the soluble fraction, (2) following the addition of a molar equivalent of GTP to Gt, and (3) in the presence of a molar equivalent of non-hydrolysable GTPγS to Gt. After illumination, the levels of Gt•GDP decreased in all three cases but were replenished in condition 2, the supplementary GTP experiment (Extended Data Fig. 7g–i). Linking Gt signalling with PDE6•cGMP hydrolysis, we monitored the release of the hydrolysed GMP product via the ratio PDE6:PDE6•cGMP. For dark-adapted membranes, before illumination, a PDE6:PDE6•cGMP ratio of approximately 1:1 was observed, consistent with full occupancy of one substrate-binding site in PDE6 (Fig. 3d, top). Signalling of rho* through Gt prompts further hydrolysis of cGMP with supplementary GTP (PDE6:PDE6•cGMP ratio of approximately 1:0.25) compared with endogenous levels (PDE6:PDE6•cGMP ratio of approximately 1:0.85) (Extended Data Fig. 7g, h). As Gαt•GTPγS is also able to interact with PDE6 (ref. 22), an intermediate level of hydrolysis of cGMP was observed in the presence of an equimolar aliquot of Gαt•GTPγS (PDE6:PDE6•cGMP of approximately 1:0.4) (Fig. 3d, bottom, Extended Data Fig. 7i). An additional PDE6–Gαt•GTP complex with 1:1 stoichiometry can also be discerned at low intensity in the presence of Gαt•GTPγS (Fig. 3d, inset). Under these experimental conditions, 1:2 complexes of PDE6•(Gαt•GTP)2 and PDE6•(cGMP)2 were not observed, consistent with existing mechanistic models. According to one model, although controversial23, one catalytic subunit of PDE6 binds to Gαt•GTP with high affinity but low activity for hydrolysing cGMP, whereas the other catalytic subunit of PDE6 binds to Gαt•GTP with low affinity but is able to hydrolyse cGMP at a high rate11. Our data, with only one Gαt•GTP subunit binding to PDE6 and with a high intensity for a single binding site of cGMP, are in accordance with the asymmetry of the activated complex and its ability to hydrolyse one cGMP to GMP per PDE6 tetramer. This reaction in turn is regulated by available GTP, activation of rho* through Gt and causes decreased concentrations of cGMP for closure of the ion channel.
Having established a means of capturing the entire signalling cascade we considered further the influence of the membrane through the effect of lipid microenvironments on rho* signalling through Gt. We examined lipids in the spectrum of dark-adapted rho expelled from membranes in the presence of Gt and PDE6 (see Methods) and found a predominance of long-chain polyunsaturated phosphatidylcholinelipids (PC 40:6, 40:8, 38:6, 38:8, 34:1, 34:2) (Extended Data Fig. 8a) in line with observations of distinct regions of retinal rods in nanodisc experiments7. After exposure to light (6–18 s) in the presence of Gt and PDE6, we observed a notable increase in the ratio of unsaturated to saturated PC lipids compared with initial and later time points (Extended Data Fig. 8a, b). Initial contact with unsaturated lipids is in line with findings of a molecular dynamic simulation study of a GPCR in an inactive or partially active state receptor24 and NMR experiments25. The increase of unsaturated lipid chains associated to rho during light activation provides evidence to support earlier proposals24 that polyunsaturated chains associate specifically with rho* to enable its conformational change from its ground state and thereby facilitate swift signalling.
Monitoring signalling in a membrane environment provides us with a unique opportunity to probe the effects of rho-targeting molecules, identified through cell-based assays designed to select compounds that enhance or perturb rho dimerization26,27 (see Methods). We tested nine compounds individually and measured their effect on rho khyd against a control (compounds 1–9; Extended Data Fig. 9a, b). The results allowed these ligands to be divided into two groups that were capable of either accelerating with a marginal effect or, more commonly, slowing retinal hydrolysis. Compounds 1 and 6 represent these two capabilities and were selected for further study (Fig. 4a). Clear differences emerge post-illumination (3 min) as rho and opsin predominate in the presence of 1 and 6, respectively (Fig. 4b, c). After 15 min, noticeable differences occur in the rho to opsin conversion, which is faster when modulated by 1 than by 6, with 1 being marginally faster than the control. As we see no evidence for displacement of retinal by these compounds or changes in the conformation of rhodopsin27, we speculate that 1 and 6 act as allosteric modulators. Calculating rates of hydrolysis and isomerization in the presence of hydroxylamine, we found that 1 accelerates both hydrolysis and isomerization, whereas rho* bound to 6 has an accelerated isomerization rate but retains retinal by decelerating hydrolysis, thereby potentially maintaining active signalling states for extended periods.
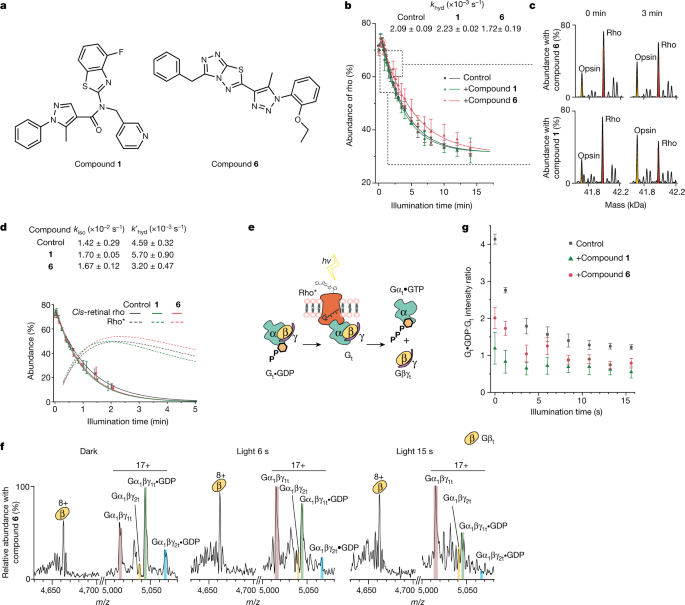
a, Structures of the two rho-targeting compounds (1 and 6) used here. b, Monitoring the conversion of rho to opsin via the change in mass following illumination in the presence of 1 or 6 and in a control under the same conditions. Data are presented as mean ± s.e. (n = 3). c, Time points for spectra of rho (red) and opsin (yellow) ejected from membranes at 0 and 3 min in the presence of 1 (bottom) and 6 (top) are shown on a zero-charge scale. d, Abundance of cis-retinal rho and rho* as a function of time in the absence (grey) or presence of 6 (pink) or 1 (green). The function for cis-retinal rho is derived from fitting data from more than 3.6 s (dashed lines) and the function for rho* is derived from the fit of the data from 18 s to 300 s (solid lines) and extrapolated (dotted lines). Data are presented as mean ± s.e. (n = 3). e, Reaction scheme: following activation with light, rho* signals through Gt to the nucleotide-free form (Gt), which then dissociates to form βγ and α. f, Mass spectra of Gt in the presence of compound 6 added to the native membrane in the absence of light (dark), and after exposure to light for 6 s and 15 s. g, Monitoring signalling through Gt via changes in the ratio of Gt•GDP:Gt in the presence or absence of compound 1 or compound 6 as a function of illumination time. Data are presented as mean ± s.e. (n = 3).
Anticipating that the prolonged activation state of rho* in the presence of 6, and the rapid isomerization and hydrolysis of 1, would affect signalling differently through Gt, we established a further assay in which we measured the ability of 1 and 6 to affect phototransduction. As above, we monitored their impact by measuring the change in mass of the conversion of Gt•GDP to apo-Gt. Before illumination, in the presence of 1 or 6 and a control under dark conditions, the ratio of Gt•GDP:Gt was 1.2, 2.0 and 4.1, respectively. In the presence of the ROS membrane, both ligands had caused significant conversion of Gt•GDP:Gt compared with the control (Fig. 4f, Extended Data Fig. 10). Unexpectedly, 1 and 6 induced transduction in the dark, implying that their enhanced isomerization rates enable them to signal through the initial trans-retinal rho population before illumination. Once illuminated, phototransduction proceeded further (Gt•GDP:Gt ratios of 0.6, 0.8 and 1.2 after 15 s for 1, 6 and the control, respectively; Fig. 4g). We conclude that 1 and 6 target rho and amplify signalling through Gt, making them ideal starting points for further chemical development. More generally, the fact that we can separate these reactions into isomerization, hydrolysis and influence on a downstream effector enables us to study rho-targeting compounds in molecular detail via their effects on phototransduction, in real time and within their native membrane environments.
Together, our native mass spectrometry data document that we have conserved a signalling cascade for an archetypal class A GPCR across its native membrane. We have shown that all three key players in the cascade can be ejected simultaneously from vesicles formed from native ROS disc membranes. In so doing, we have captured the conversion of rho to opsin in real time and have shown that it is significantly slower in the membrane than in the detergent micelle. We observed regeneration of rho following exposure to light and detected changes in the level of N-ret-PE and depletion of the conjugated chromophore following illumination. The importance of the membrane also prompted us to consider the changing lipid microenvironment during signalling and documented changes in the association of unsaturated long-chain PC lipids in the vicinity of signalling rho. Considering the effects of rho-targeting compounds, we demonstrated their ability to accelerate the rho to opsin conversion or slow down the reaction, and we hypothesize that the latter would stabilize a signalling-competent state. In line with this hypothesis, we demonstrated the ability to modulate the Gt signalling pathway through an increase in the turnover of Gt•GDP to Gt and the subsequent dissociation to the Gβγt complex and Gαt•GTP primed for interaction with PDE6. We observed the static intermediate Gαt–PDE6, as well as the effects of endogenous GTP levels on the hydrolysis of cGMP through light-activated signalling across the membrane. By capturing the entire signalling process, we have demonstrated the importance of the lipid microenvironment for signalling, coupling, effector targeting and regeneration. We anticipate that similar approaches will be applicable to other cell-surface receptors (for example, olfactory receptors) present at high density, enabling kinetic studies of molecular events, and identification of endogenous and synthetic receptor ligands that perturb signalling. Overall, we have highlighted an approach to drug discovery in which different stages of a signalling cascade can be targeted across native membranes.
More News
I make 3D models of conifer needles to explore their climate effects
How rich is too rich?
China’s Chang’e-6 launches successfully — what happens next?