



Human-modified forests, such as selectively logged forests, are often characterized as degraded ecosystems because of their altered structure and low biomass. The concept of ecosystem degradation can be a double-edged sword. It rightly draws attention to the conservation value of old-growth systems and the importance of ecosystem restoration. However, it can also suggest that human-modified ecosystems are of low ecological value and therefore, in some cases, suitable for conversion to agriculture (such as oil palm plantations) and other land uses3,4,5.
Selectively logged and other forms of structurally altered forests are becoming the prevailing vegetation cover in much of the tropical forest biome2. Such disturbance frequently leads to a decline in old-growth specialist species1, and also in non-specialist species in some contexts6,7,8. However, species-focused biodiversity metrics are only one measure of ecosystem vitality and functionality, and rarely consider the collective role that suites of species play in maintaining ecological functions9.
An alternative approach is to focus on the energetics of key taxonomic groups, and the number and relative dominance of species contributing to each energetic pathway. Energetic approaches to examining ecosystem structure and function have a long history in ecosystem ecology10. Virtually all ecosystems are powered by a cascade of captured sunlight through an array of autotroph tissues and into hierarchical assemblages of herbivores, carnivores and detritivores. Energetic approaches shine light on the relative significance of energy flows among key taxa and provide insight into the processes that shape biodiversity and ecosystem function. The common currency of energy enables diverse guilds and taxa to be compared in a unified and physically meaningful manner: dominant energetic pathways can be identified, and the resilience of each pathway to the loss of individual species can be assessed. Quantitative links can then be made between animal communities and the plant-based ecosystem productivity on which they depend. The magnitude of energetic pathways in particular animal groups can often be indicators of key associated ecosystem processes, such as nutrient cycling, seed dispersal and pollination, or trophic factors such as intensity of predation pressure or availability of resource supply, all unified under the common metric of energy flux11,12.
Energetics approaches have rarely been applied in biodiverse tropical ecosystems because of the range of observations they require11,12,13. Such analyses rely on: population density estimates for a very large number of species; understanding of the diet and feeding behaviour of the species; and reliable estimation of net primary productivity (NPP). Here we take advantage of uniquely rich datasets to apply an energetics lens to examine and quantify aspects of the ecological function and vitality of habitats in Sabah, Malaysia, that comprise old-growth forests, logged forest and oil palm plantation (Fig. 1 and Extended Data Fig. 1). Our approach is to calculate the short-term equilibrium production or consumption rates of food energy by specific species, guilds or taxonomic groups. We focus on three taxonomic groups (plants, birds and mammals) that are frequently used indicators of biodiversity and are relatively well understood ecologically.
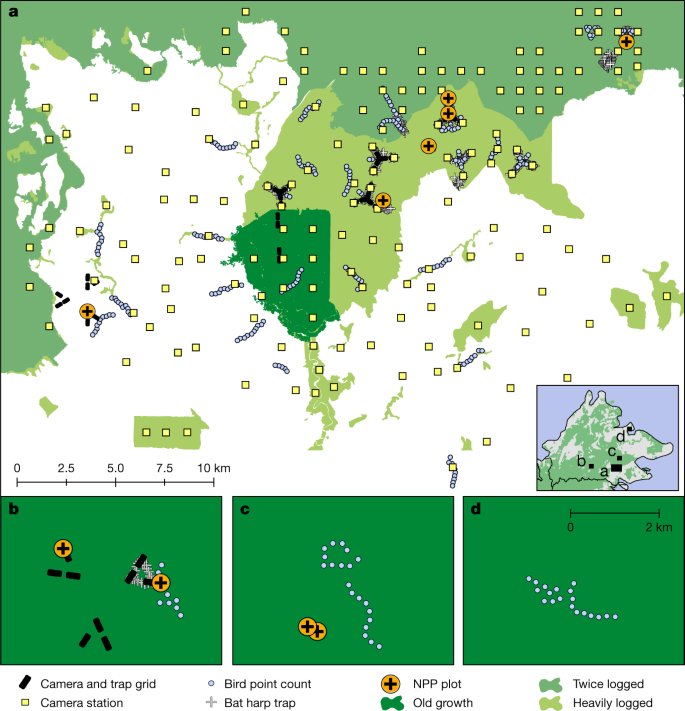
a–d, Maps showing locations of NPP plots and biodiversity surveys in old-growth forest, logged forest and oil palm plantations in the Stability of Altered Forest Ecosystems Project landscape (a), Maliau Basin (b), Danum Valley (c) and Sepilok (d). The inset in a shows the location of the four sites in Sabah. The shade of green indicates old-growth (dark green), twice-logged (intermediate green) or heavily logged (light green) forests. The camera and trap grid includes cameras and small mammal traps. White areas indicate oil palm plantations.
We are interested in the fraction of primary productivity consumed by birds and mammals, and how it varies along the disturbance gradient, and how and why various food energetic pathways in mammals and birds, and the diversity of species contributing to those pathways, vary along the disturbance gradient. To estimate the density of 104 mammal and 144 bird species in each of the three habitat types, we aggregated data from 882 camera sampling locations (a total of 42,877 camera trap nights), 508 bird point count locations, 1,488 small terrestrial mammal trap locations (34,058 live-trap nights) and 336 bat trap locations (Fig. 1 and Extended Data Fig. 1). We then calculated daily energetic expenditure for each species based on their body mass, assigned each species to a dietary group and calculated total food consumption in energy units. For primary productivity, we relied on 34 plot-years (summation of plots multiplied by the number of years each plot is monitored) of measurements of the key components of NPP (canopy litterfall, woody growth, fine root production) using the protocols of the Global Ecosystem Monitoring Network14,15,16 across old-growth (n = 4), logged (n = 5) and oil palm (n = 1) plots. This dataset encompasses more than 14,000 measurements of litterfall, 20,000 tree diameter measurements and 2,700 fine root samples.
Overall bird species diversity is maintained across the disturbance gradient and peaks in the logged forest; for mammals, there is also a slight increase in the logged forest, followed by rapid decline in the oil palm (Fig. 2b,c). Strikingly, both bird and mammal biomass increases substantially (144% and 231%, respectively) in the logged forest compared to the old-growth forest, with mammals contributing about 75% of total (bird plus mammal) biomass in both habitat types (Fig. 2b,c).
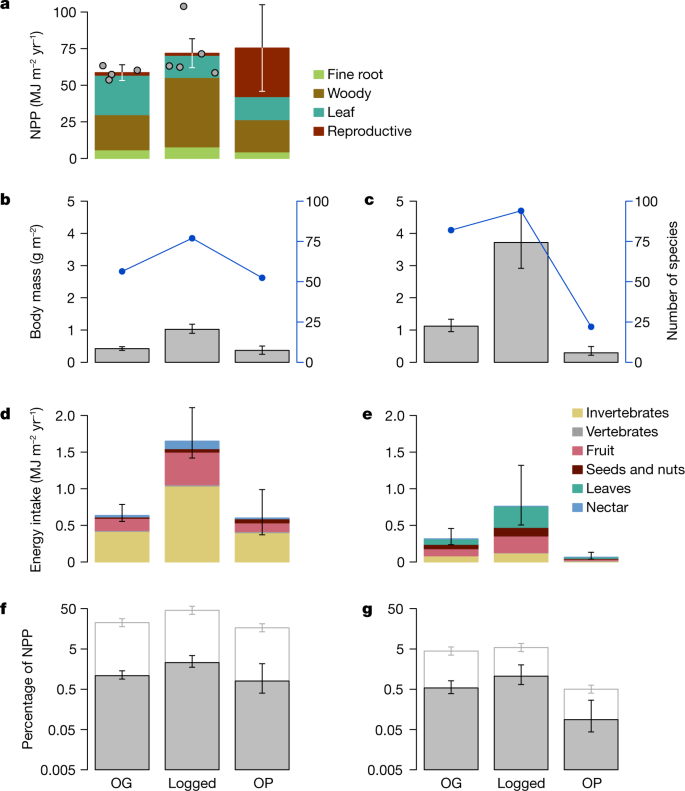
a, Total NPP along the gradient (mean of intensive 1-ha plots; n = 4 for old growth (OG), n = 5 for logged and n = 1 for oil palm (OP); error bars are 95% confidence intervals derived from propagated uncertainty in the individually measured NPP components), with individual plot data points overlaid. b,c, Total body mass (bars, left axis) and number of species counted (blue dots and line, right axis) of birds (b) and mammals (c). d,e, Total direct energetic food intake by birds (d) and mammals (e). f,g, Percentage of NPP directly consumed by birds (f) and mammals (g). In b–e, body mass and energetics were estimated for individual bird and mammal species, with the bars showing the sum. Error bars denote 95% confidence intervals derived from 10,000 Monte Carlo simulation estimates incorporating uncertainty in body mass, population density, the daily energy expenditure equation, assimilation efficiency of the different food types, composition of the diet of each species and NPP. In f,g, the grey bars indicate direct consumption of NPP, white bars denote the percentage of NPP indirectly supporting bird and mammal food intake when the mean trophic level of consumed invertebrates is assumed to be 2.5, with the error bars denoting assumed mean trophic levels of 2.4 and 2.6. Note the log scale of the y axis in f,g. Numbers for d,e provided in Supplementary Data Tables 1, 2.
The total flow of energy through consumption is amplified across all energetic pathways by a factor of 2.5 (2.2–3.0; all ranges reported are 95% confidence intervals) in logged forest relative to old-growth forest. In all three habitat types, total energy intake by birds is much greater than by mammals (Fig. 2d,e and Extended Data Table 1). Birds account for 67%, 68% and 90% of the total direct consumption by birds and mammals combined in old-growth forests, logged forests and oil palm, respectively. Although mammal biomass is higher than bird biomass in the old-growth and logged forests, the metabolism per unit mass is much higher in birds because of their small body size; hence, in terms of the energetics and consumption rates, the bird community dominates. The total energy intake by birds alone increases by a factor of 2.6 (2.1–3.2) in the logged forest relative to old-growth forest. This is mainly driven by a 2.5-fold (1.7–2.8) increase in foliage-gleaning insectivory (the dominant energetic pathway), and most other feeding guilds also show an even larger increase (Figs. 2d and 3). However, total bird energy intake in the oil palm drops back to levels similar to those in the old-growth forest, with a collapse in multiple guilds. For mammals, there is a similar 2.4-fold (1.9–3.2) increase in total consumption when going from old-growth to logged forest, but this declines sharply in oil palm plantation. Most notable is the 5.7-fold (3.2–10.2) increase in the importance of terrestrial mammal herbivores in the logged relative to old-growth forests. All four individual old-growth forest sites show consistently lower bird and mammal energetics than the logged forests (Extended Data Fig. 5).
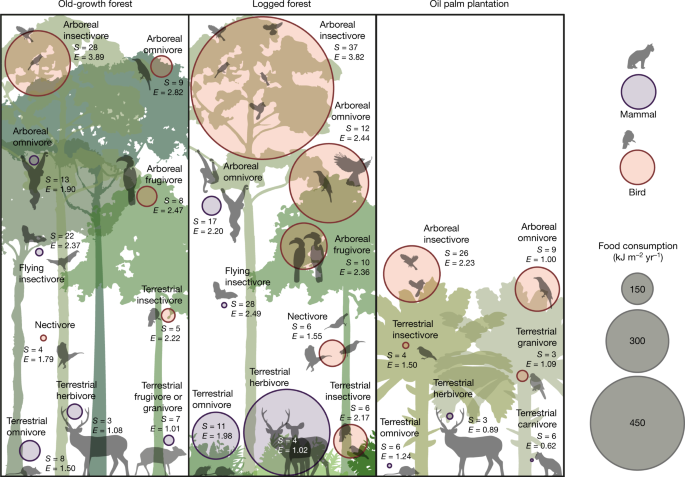
The size of the circles indicates the magnitude of energy flow, and the colour indicates birds or mammals. S, number of species; E, ESWI, an index of species redundancy and, therefore, resilience (high values indicate high redundancy; see main text). For clarity, guilds with small energetic flows are not shown, but are listed in Supplementary Data 4. Images created by J. Bentley.
The fraction of NPP flowing through the bird and mammal communities increases by a factor of 2.1 (1.5–3.0) in logged forest relative to old-growth forest. There is very little increase in NPP in logged relative to old-growth forests (Fig. 2a) because increased NPP in patches of relatively intact logged forest is offset by very low productivity in more structurally degraded areas such as former logging platforms14,15. In oil palm plantations, oil palm fruits account for a large proportion of NPP, although a large fraction of these is harvested and removed from the ecosystem17. As a proportion of NPP, 1.62% (1.35–2.13%) is directly consumed by birds and mammals in the old-growth forest; this rises to 3.36% (2.57–5.07%) in the logged forest but drops to 0.89% (0.57–1.44%) in oil palm (Fig. 2f,g and Extended Data Table 2).
If all invertebrates consumed are herbivores or detritivores (that is, at a trophic level of 2.0), and trophic efficiency is 10% (ref. 10), the total amount of NPP supporting the combined bird and mammal food intake would be 9%, 16% and 5% for old-growth forest, logged forest and oil palm, respectively. However, if the mean trophic level of consumed invertebrates is 2.5 (that is, a mix of herbivores and predators), the corresponding proportions would be 27%, 51% and 17% (Fig. 2f,g). As insectivory is the dominant feeding mode for the avian community, these numbers are dominated by bird diets. For birds in the old-growth forests, 0.35% of NPP supports direct herbivory and frugivory, but around 22% of NPP (assumed invertebrate trophic level 2.5) is indirectly required to support insectivory. The equivalent numbers for birds in logged forest are 0.83% and 46%. Hence, birds account for a much larger indirect consumption of NPP. Bird diet studies in old-growth and logged forest in the region suggest that consumed invertebrates have a mean trophic level of 2.5 (ref. 18; K. Sam, personal communication), indicating that the higher-end estimates of indirect NPP consumption (that is, around 50% in logged forests) are plausible.
It is interesting to compare such high fractions of NPP to direct estimates of invertebrate herbivory. Scans of tree leaf litter from these forests suggest that just 7.0% of tree canopy leaf area (1–3% of total NPP) is removed by tree leaf herbivory14,16, but such estimates do not include other pathways available to invertebrates, including herbivory of the understorey, aboveground and belowground sap-sucking, leaf-mining, fruit- and wood-feeding, and canopy, litter and ground-layer detritivory. An increase in invertebrate biomass and herbivory in logged forest compared to old-growth forest has previously been reported in fogging studies in this landscape19. Such high levels of consumption of NPP by invertebrates could have implications on ecosystem vegetation biomass production, suggesting, first, that invertebrate herbivory has a substantial influence on recovery from logging and, second, that insectivorous bird densities may exert substantial indirect controls on ecosystem recovery.
The distributions of energy flows among feeding guilds are remarkably stable among habitat types (Fig. 3), indicating that the amplified energy flows in the logged forests do not distort the overall trophic structure of vertebrate communities. Overall bird diet energetics are dominated by insectivory, which accounts for a strikingly invariant 66%, 63% and 66% of bird energetic consumption in old-growth forest, logged forest and oil palm, respectively. Foliage-gleaning dominates as a mode of invertebrate consumption in all three habitat types, with frugivory being the second most energetically important feeding mode (26%, 27% and 19%, respectively). Mammal diet is more evenly distributed across feeding guilds, but frugivory (31%, 30%, 30%) and folivory (24%, 38%, 26%) dominate. Small mammal insectivores are probably under-sampled (see Methods) so the contribution of mammal insectivory may be slightly greater than that estimated here. The apparent constancy of relative magnitude of feeding pathways across the intact and disturbed ecosystems is noteworthy and not sensitive to plausible shifts in feeding behaviour between habitat types (see Supplementary Discussion). There is no evidence of a substantial shift in dominant feeding guild: the principal feeding pathways present in the old-growth forest are maintained in the logged forest.
When examining change at species level in the logged forests, the largest absolute increases in bird food consumption were in arboreal insectivores and omnivores (Fig. 4a and Extended Data Fig. 2a). In particular, this change was characterized by large increases in the abundance of bulbul species (Pycnonotus spp.). No bird species showed a significant or substantial reduction in overall energy consumption. In the oil palm plantation, total food consumption by birds was less than in logged forests, but similar to that in old-growth forests. However, this was driven by very high abundance of a handful of species, notably a single arboreal omnivore (yellow-vented bulbul Pycnonotus goiavier) and three arboreal insectivores (Mixornis bornensis, Rhipidura javanica, Copsychus saularis), whereas energy flows through most other bird species were greatly reduced (Fig. 4b and Extended Data Fig. 2b).
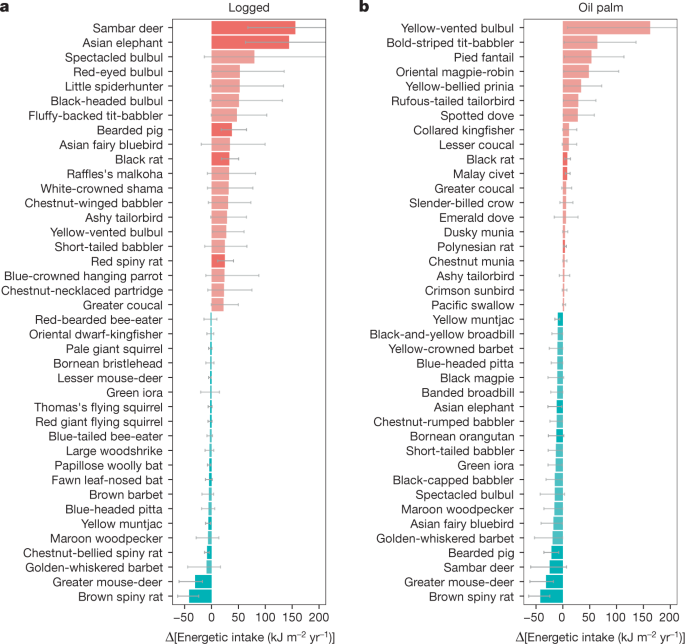
a,b, Changes in energy consumption by species in logged forest relative to old-growth forest (a) and in oil palm relative to old-growth forest (b). The 20 species experiencing the largest increase (red) and decrease (blue) in both habitat types are shown. Bird species are shown in a lighter tone and mammal species are shown in a darker tone. The error bars denote 95% confidence intervals, derived from 10,000 Monte Carlo simulation estimates incorporating uncertainty in body mass, population density, the daily energy expenditure equation, assimilation efficiency of the different food types and composition of the diet of each species.
For mammals, the increase in consumption in logged forests is dominated by consumption by large terrestrial herbivores increasing by a factor of 5.7 (3.2–10.2), particularly sambar deer (Rusa unicolor) and Asian elephant (Elephas maximus; Fig. 4a and Extended Data Figs. 2b and 3), along with that by small omnivores, predominantly rodents (native spiny rats, non-native black rat; Fig. 4). A few rainforest species show a strong decline (for example, greater mouse-deer Tragulus napu and brown spiny rat Maxomys rajah). In the oil palm, most mammal species collapse (Fig. 4b) and the limited consumption is dominated by a few disturbance-tolerant habitat generalists (for example, red muntjac Muntiacus muntjak, black rat Rattus rattus, civets), albeit these species are at lower densities than observed in old-growth forest (Extended Data Fig. 2).
With very few exceptions, the amplified energy flows in logged forest seem to retain the same level of resilience as in old-growth forest. The diversity and dominance of species within any pathway can be a measure of the resilience of that pathway to loss of species. We assessed energetic dominance within individual pathways by defining an energetic Shannon–Wiener index (ESWI) to examine distribution of energy flow across species; low ESWI indicates a pathway with high dependence on a few species and hence potential vulnerability (Fig. 3). The overall ESWI across guilds does not differ between the old-growth and logged forest (t2,34 = −0.363, P = 0.930), but does decline substantially from old-growth forest to oil palm (t2,34 = −3.826, P = 0.0015), and from logged forest to oil palm (t2,34 = −3.639, P = 0.0025; linear mixed-effects models, with habitat type as fixed effect and guild as random effect; for model coefficients see Supplementary Table 3).
Hence, for birds, the diversity of species contributing to dominant energetic pathways is maintained in the transition from old-growth to logged forests but declines substantially in oil palm. Mammals generally show lower diversity and ESWI than birds, but six out of ten feeding guilds maintain or increase ESWI in logged forest relative to the old-growth forests but collapse in oil palm (Fig. 3). Terrestrial herbivory is the largest mammal pathway in the logged forest but is dependent on only four species and is probably the most vulnerable of the larger pathways: a few large mammals (especially sambar deer) play a dominant terrestrial herbivory role in the logged forest. In parallel, bearded pigs (Sus barbatus), the only wild suid in Borneo, form an important and functionally unique component of the terrestrial omnivory pathway. These larger animals are particularly sensitive to anthropogenic pressures such as hunting, or associated pathogenic pressures as evidenced by the recent precipitous decline of the bearded pig in Sabah due to an outbreak of Asian swine fever (after our data were collected)20.
Vertebrate populations across the tropics are particularly sensitive to hunting pressure21. Our study site has little hunting, but as a sensitivity analysis we explored the energetic consequences of 50% reduction in population density of those species potentially affected by targeted and/or indiscriminate hunting (Extended Data Fig. 4). Targeted hunted species include commercially valuable birds, and gun-hunted mammals (bearded pig, ungulates, banteng and mammals with medicinal value). Indiscriminately hunted species include birds and mammals likely to be trapped with nets and snares. Hunting in the logged forests lowers both bird and mammal energy flows but still leaves them at levels higher than in faunally intact old-growth forests. Such hunting brings bird energetics levels close to (but still above) those of old-growth forests. For mammals, however, even intensively hunted logged forests seem to maintain higher energetic flows than the old-growth forests. Hence, only very heavy hunting is likely to ‘offset’ the amplified energetics in the logged forest.
The amplified energetic pathways in our logged forest probably arise as a result of bottom-up trophic factors including increased resource supply, palatability and accessibility. The more open forest structure in logged forest results in more vegetation being near ground level22,23 and hence more accessible to large generalist mammal herbivores, which show the most striking increase of the mammal guilds. The increased prioritization by plants of competition for light and therefore rapid vegetation growth strategies in logged forests results in higher leaf nutrient content and reduced leaf chemical defences against herbivory24,25, along with higher fruiting and flowering rates19 and greater clumping in resource supply9. This increased resource availability and palatability probably supports high invertebrate and vertebrate herbivore densities25. The act of disturbance displaces the ecosystem from a conservative chemically defended state to a more dynamic state with amplified energy and nutrient flow, but not to an extent that causes heavy disruption in animal community composition. Top-down trophic factors might also play a role in amplifying the energy flows in intermediate trophic levels, through mechanisms such as increased protection of ground-dwelling or nesting mammals and birds from aerial predators in the dense vegetation ground layer. This might partially explain the increased abundance of rodents, but there is little evidence of trophic release at this site because of the persisting high density of mammal carnivores26. Overall, the larger number of bottom-up mechanisms and surge in invertebrate consumption suggest that increased resource supply and palatability largely explains the amplification of consumption pathways in the logged forest. An alternative possibility is that the amplified vertebrate energetics do not indicate amplified overall animal energetics but rather a large diversion of energy from unmeasured invertebrate predation pathways (for example, parasitoids); this seems unlikely but warrants further exploration.
Oil palm plantations show a large decline in the proportion of NPP consumed by mammals and birds compared to logged forests12. Mammal populations collapse because they are more vulnerable and avoid humans, and there is no suite of mammal generalists that can step in27,28. Birds show a more modest decline, to levels similar to those observed in old-growth forests, as there is a broad suite of generalist species that are able to adapt to and exploit the habitat types across the disturbance gradient, and because their small size and mobility render them less sensitive to human activity29. There is a consistent decline in the oil palm in ESWI for birds and especially for mammals, indicating a substantial increase in ecosystem vulnerability in many pathways.
In conclusion, our analysis demonstrates the tremendously dynamic and ecologically vibrant nature of the studied logged forests, even heavily and repeatedly logged forests such as those found across Borneo. It is likely that the patterns, mechanisms and basic ecological energetics we describe are general to most tropical forests; amplification of multiple ecosystem processes after logging has also been reported for logged forests in Kenya9, but similar detailed analyses are needed for a range of tropical forests to elucidate the importance of biogeographic, climatic or other factors. We stress that our findings do not diminish the importance of protecting structurally intact old-growth forests, but rather question the meaning of degradation by shining a new light on the ecological value of logged and other structurally ‘degraded’ forests, reinforcing their significance to the conservation agenda30. We have shown that a wide diversity of species not only persist but thrive in the logged forest environment. Moreover, such ecological vibrancy probably enhances the prospects for ecosystem structural recovery. In terms of faunal intactness, our study landscape is close to a best-case scenario because hunting pressures were low. If logged forests can be protected from heavy defaunation, our analysis demonstrates that they can be vibrant ecosystems, providing many key ecosystem functions at levels much higher than in old-growth forests. Conservation of logged forest landscapes has an essential role to play in the in the protection of global biodiversity and biosphere function.
More News
Japan can embrace open science — but flexible approaches are key
Beware of graphene’s huge and hidden environmental costs
Countering extreme wildfires with prescribed burning can be counterproductive